
Leishmania is a protozoan parasite responsible for leishmaniasis, a vector-borne disease that causes painful and often disfiguring tissue destruction. Leishmania major is known for causing cutaneous leishmaniasis and is mainly found in the Middle East and North Africa. Leishmania donovani causes visceral leishmaniasis, where the tissue destruction occurs in the organs. This manifestation is primarily known to occur in India and East Asia.
In order to complete its lifecycle, Leishmania undergoes two vital development stages. One stage is in a mammalian (e.g. human) host and the other in the sand fly digestive system. Leishmania survival in the sand fly host is an important stage to investigate as different sand fly species vary in their ability to transmit the disease (approximately 30 species of sand fly can spread at least 20 species of Leishmania). This measure of transmission ability is called vector competence and understanding the factors that affect it can aid attempts to eradicate this often-deadly disease.
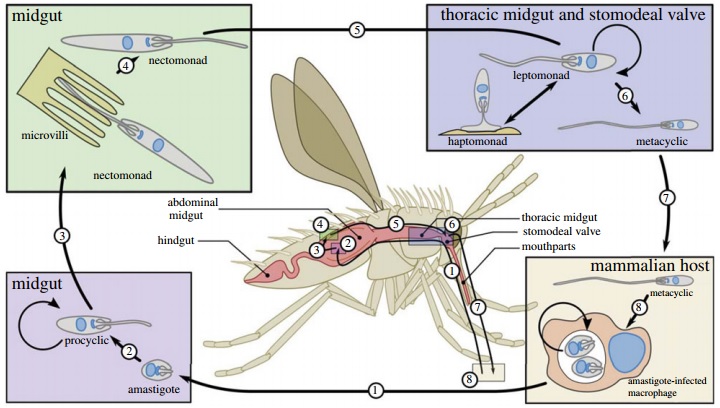
Lifecycle in the sand fly
The sand fly feeds on blood from an infected mammal and takes up the amastigote form of the parasite into the abdominal midgut. Here the parasite transforms to the procyclic promastigote form. The parasite is then thought to attach to the midgut wall and finally migrate back towards the mouthparts. Just before it reaches the mouthparts it converts to its mammalian-infectious form, the metacyclic promastigote, ready for when the sand fly takes its next blood meal. The parasite is passed on because the sand fly regurgitates the mixture that lingers in its mouth parts into the bite before feeding.
Pruzinova and colleagues looked into parasite developmental success in two competent and two non-competent (refractory) sand fly species and found some surprising results. The first part of their study consisted of in vitro experiments and can be broken down into 3 main findings (although more aspects can be found in the paper itself):
- death in the midgut is not sand fly-species-specific
- time since blood meal affects midgut toxicity
- midgut proteases are likely not the direct cause of death.
The second part of the study looks at how the in vitro findings relate to their in vivo findings.
Death by midgut is not sand fly species-specific
This experiment investigates whether the midgut contents of different sand fly species have varying levels of toxicity to L. donovani and L. major. The authors dissected out midguts from four sand fly species and mixed them with the parasites. Surprisingly, the midguts of the two refractory sand fly hosts did not cause higher mortality than the two competent sand fly species.
Time since blood meal affects midgut toxicity
Again, midguts were dissected out from four sand fly species, but at different time intervals since feeding, and incubated with parasites. All sand fly species midguts were most toxic towards the end of the process of digestion of their blood meal. After analysing these data, the authors concluded that parasite death in these in vitro experiments is not so much dependent on sand fly species, but rather, dependent on time since the sand fly last fed.
Midgut proteases are likely not the direct cause of death
Previous explanations for parasite death in refractory flies have included the hypothesis that sand fly proteases are responsible. This study incubated L. donovani parasites with rabbit blood or human haemoglobin and/or with a commercially bought proteinase. Interestingly the parasites were killed by blood/haemoglobin and the proteinase together, but not by the proteinase on its own. The authors argue that, instead of previous opinion, it could be toxins produced during blood digestion (e.g haem) that are harmful to the parasites rather than the digestion agents (e.g. proteases) themselves. This suggestion fits with the above finding where midgut contents were more toxic the longer the sand fly had been digesting.
How do these findings relate to the in vivo environment?
Excitingly, this group also conducted an in vivo study where the two susceptible and two refractory sand fly species were infected with L. donovani. Parasite development was examined by dissecting out each section of the gut at different times post-blood meal and identifying the parasite life stage. An additional explanation that has been suggested for parasite mortality in the sand fly Phlebotomus papatasi is parasite inability to attach to the gut wall. The data here show that the parasites do survive and develop in P. papatasi but disappear after sand fly defecation. Therefore, it seems in this in vivo scenario that L. donovani lose grip in the gut rather than being killed by toxins.
A grip mechanism has been described as the strategy that enables L. major to be successful in P. papatasi, whereby parasite lipophosphoglycan (LPG) binds to sand fly galectins. The results from this study by Pruzinova and colleagues therefore support the idea that L. donovani do not possess the appropriate LPG for binding.
Conclusion
This study looked at several aspects of Leishmania development within different sand fly species and found that any death caused by the contents of the midgut, specifically proteases, was not significantly species-specific but dependent on the stage of blood digestion. In addition, as seen by the in vivo study, L. donovani can initially survive in a refractory sand fly species but are eventually lost through defecation.
Comments