Caste formation and how it works
The allocation of specific jobs to individuals and the co-operative division of labour within a colony is a well-established phenomenon called a ‘caste system’, and has been thoroughly characterised within organisms with hierarchical communities, such as ants and bees. Caste formation has also been recognised in reef-shrimp, sea anemones and mole-rats.
However, in recent years there has been a growing body of evidence showing that a complex caste system is present in a number of digenean trematodes (parasitic worms).
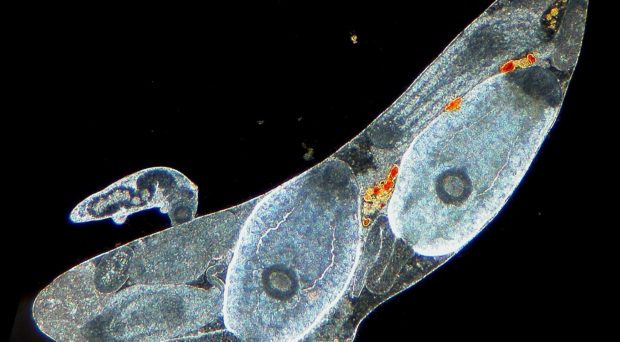
In a tightly organised society with a caste system, individuals are often divided into those that are reproductive and non-reproductive. Those that are non-reproductive (soldiers) are morphologically distinct from their reproductive counterparts and are specialised to defend the colony from invasion by attacking and killing invading organisms.
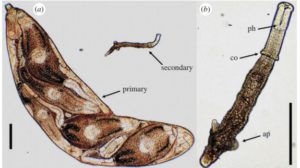
In trematodes, the caste system comes into effect during the colonial stage of its lifecycle, when it inhabits an aquatic snail. A miracidium hatches from an egg deposited in the water by its definitive host. The miracidium then locates and infects its intermediate snail host, and if successful, establishes a period of proliferative asexual development (a colony), thereby producing >10,000 genetically identical larvae. Larval trematodes (rediae) allocated as soldiers are smaller and thinner than their reproductive colleagues, but have relatively large mouthparts and have been shown to more active (and disproportionately deployed) in areas of the snail host where invasion by other trematode colonies can occur.
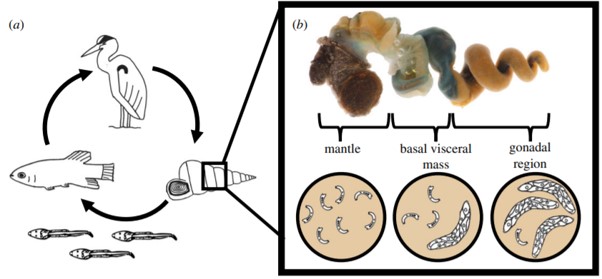
Studies in 2017 and 2020 have both utilised Cerithideopsis californic to research the caste formation phenomena. The California horn snail is a marine gastropod recognised to play host to a diverse range of trematode species (at least 18), and so provided a perfect organism to research colony competition and the mechanisms behind soldier allocation.
Standing army tax and the optimal allocation theory
The mechanism by which larval trematodes are allocated as either soldiers or reproductives is currently undetermined. However, it is known that resources are limited, and defences are costly. Much like the medieval kingdoms of Europe, trematodes pay a heavy cost to keep a standing army ready to respond to an invasion. However, the cost they pay for an army isn’t monetary like the feudal lords of old but is instead paid in the form of decreased reproductive output due to allocation of larvae as soldiers, rather than their reproducing forms.

Emlyn Resetarits and her colleagues theorised that allocating more of the colony to reproduction should optimise the colonies fitness (i.e. success) at low invasion threat levels, but that when the colony is “exposed to higher threat levels, allocating more to defence should be favoured.” This theory is known as the optimal allocation theory, and it predicts that within a society with a soldier caste, allocation to soldiers should essentially reflect the level of threat of invasion.
Two hypotheses of optimal soldier allocation have been presented: inducible and constitutive mechanisms. Inducible (plastic) allocation infers that there will be different numbers of soldiers allocated to areas under a high threat of invasion (invasion fronts) in real time, whereas constitutive (innate) allocation essentially says that soldier allocation is fixed in a colony overtime, and that an increased threat of invasion in a specific area of the snail host doesn’t promote increased soldier allocation.
Despite indications that ants, wasps, and even a species of trematode can increase the numbers of soldiers in response to an increase in threat, we currently lack strong evidence if differences in the location of that threat can in some way influences soldier allocation.
A plastic mechanism of soldier allocation when threat levels are high has indeed been shown in social insects and a species of trematode, however it has only been shown in trematodes when actively waring with an invading trematode of a different species, otherwise known as a heterospecific invader.
Therefore, Emlyn Resetarits and colleagues attempted to quantify invasion threat and solider allocation in 168 trematode colonies for six trematode species (one colony and trematode species per snail), across a range of invasion threat levels to shed light on which soldier allocation mechanism parasitic trematodes utilise.
Soldiers ready!
Results from the study detail that soldier allocation substantially increased with an increased threat of invasion, and thus supporting a plastic mechanism of soldier allocation, and the optimal allocation theory. Interestingly, all six species of trematode shared a similar response, significantly increasing their solider allocation when the perceived threat level increased.
In spite of the demonstrated increase in soldier numbers and evidence of the plasticity of soldier allocation, it appears that trematode colonies do not in fact deploy a great number of soldiers to high threat invasion fronts. Rather, they opt to simply have a larger standing army to better defend their colony, potentially signifying that these trematodes can perceive risk, perhaps by even ‘remembering’ previously failed invasion attempts.
Competitive eating
Understandably, co-infection of the same snail host by multiple trematodes (colonies or species) creates opportunities for competition. For instance, it’s been shown that larval echinostomes predate on larval schistosomes in their shared snail host, and demonstrated competition with and dominance over schistosomes species. Martina Laidemitt and colleagues have suggested that evidence that echinostomes could reduce the prevalence of schistosomes implies they could be used as a biological control agent, and could provide an attractive approach to controlling Schistosomiasis.
If you wish to read more about this fascinating topic then please look at our other blog post regarding trematode soldiers.

Comments