
Current global changes impact all biota. There are those species that can utilize the changes and those that will have a hard time coping with the human induced changes. Invaders are among the winners and pose a serious threat to native species. Mitigation of the problems associated with invaders has sparked interest in predicting which type of invader species and genotypes could become invaders. However, invasion is always an interplay of the invader’s and resident community’s characteristics. Therefore one should remember that in the changing world anthropogenic change affects not only the invader but also community composition. Moreover, different areas are known to harbour different kinds of species assemblages. It is, thus, worthwhile to ask which kinds of communities could be more vulnerable to invasions.
Let’s make it clear that this idea is by no means new. Many researchers since Charles Darwin and Charles S. Elton have done research on this subject: correlating species diversity, phylogenetic diversity, and relatedness between the community and the invader. Despite decades of work on invasions and coexistence mechanisms the evidence seems to be mixed, regardless of the study taxa and methodology. One thing, however, is rather clearly evident: more systems where key aspects of invasion theories can be manipulated are in demand, as from naturally occurring invasions it is hard to explicitly show the determinants of invasions.
In our recent publication in BMC Ecology we present a microbial system for the study of invasion theories, and use it to show that diversity in phylogenetic similarity between the invader and the community members (i.e. the closeness of their evolutionary relationships) affects invasion success. Those communities with diversity in phylogenetic distances to invaders resist invasions better than communities that are less diverse in their phylogenetic relationships to an invader.
This is actually a rather novel finding, as effects of diversity on invasions have been studied mostly at ecological level (species diversity), and not at the evolutionary level of phylogenetic diversity. Since invasion was explained by a metric that describes relatedness between the invader and the community our result casts a doubt on easy recognition of potential invaders, or vulnerable communities. Invader and community act together in the invasion process.
Blaah – microbes – you might think, close the link, and forget that lots of ecological and evolutionary theory has been created and tested with microbes. But, microbes invade as well, and knowing about microbial invasions of different habitats – lakes, pools, foods, teeth, wounds, and lungs – has direct implications for our day-to-day life.
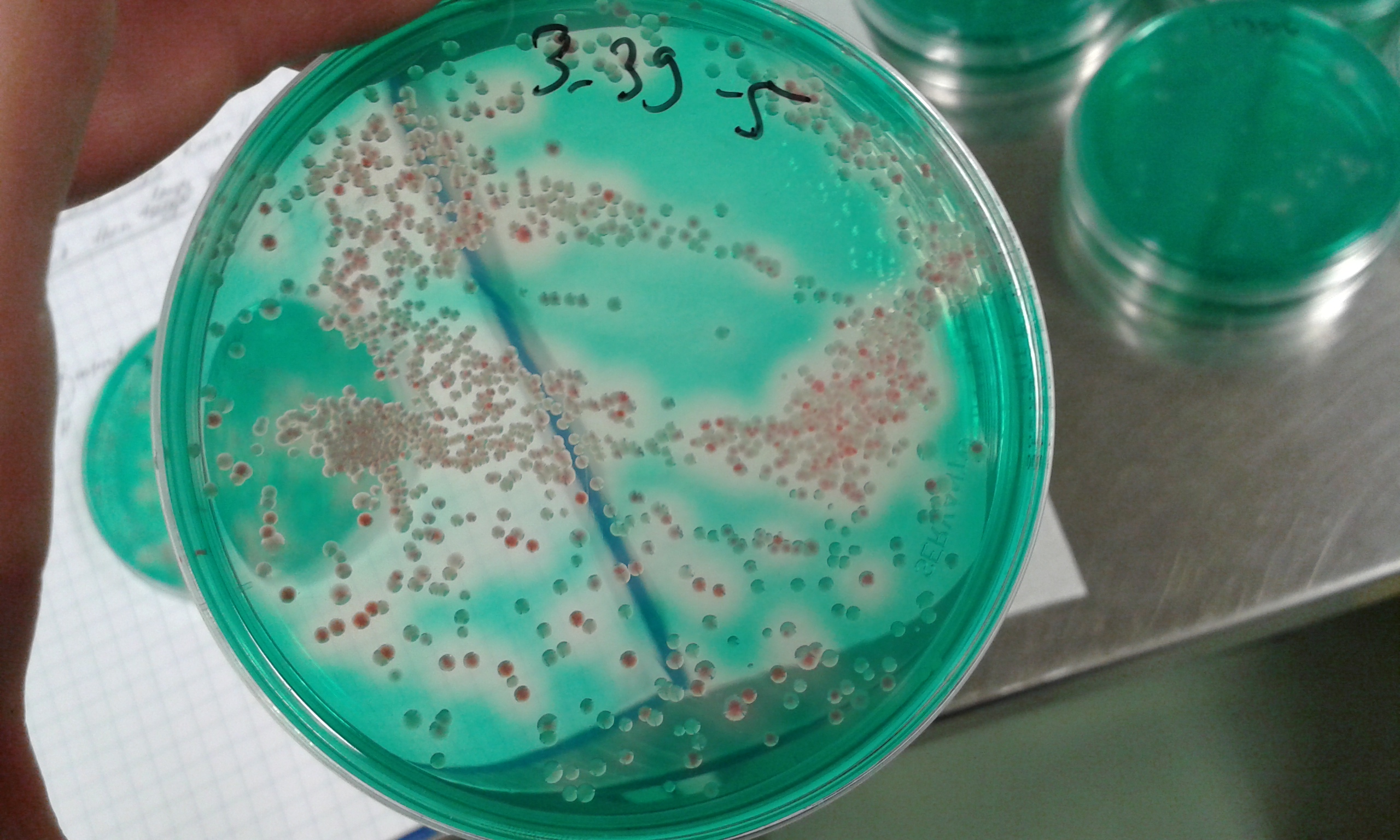
In our experiments we used the opportunistic environmentally growing pathogen Serratia marcescens as an invader. This species infects literally everything. For healthy humans, it is rather harmless, but it hosts range span from corals and plants to insects. Several species of insects, when infected, die within few days.
The coolest thing in our system is that we can identify it from other bacteria with very simple agar plating. In addition to red pigment, S. marcescens creates a halo around its colonies in DNAse agar plates, whereas other bacteria in our system are seen as white colonies without a halo. The other bacteria are traditional “lab rats” in microbiology like E. coli, and Pseudomonas species.
This system can be used to test several key hypotheses concerning invasions; not only by manipulating species in the community but also their evolutionary background. Easily culturable bacteria can be put through experimental evolution with ease. Such new adapted genotypes can be used to create different invasion scenarios. For example mimicking anthropogenic change – and this is the direction where we are going to head next.
Comments