Development of robust genetic research systems to explore the biology of Plasmodium falciparum has lagged far behind model organisms such as yeast and Drosophila. In two recent back-to-back reports in Nature, however, the application of three novel (at least for Plasmodium!) conditional expression knockdown systems are used to explore the function of the Plasmodium translocon of exported proteins (PTEX) in the modification of the human erythrocyte host during intraerythrocytic growth and division.
Intraerythrocytic development (schizogony) requires that the parasite export hundreds of proteins into its host cell to facilitate successful colonization, adaption and multiplication within its host – with several of these proteins critical for the pathophysiology of disease. As the parasite resides within a parasitophorous vacuole (PV) it must also export a protein trafficking complex into the PV membrane to ensure export into the host cytoplasm and beyond – this is the PTEX. The key components of the PTEX are known, as is the role of a Plasmodium export element (PEXEL) motif in marking proteins for export beyond the PV membrane (see figure below and a recent review article). However, the essentiality of these processes proved a significant barrier to more involved studies of function when the only molecular tool available was a straight forward knockout. The studies reported by the de Koning-Ward (Deakin University, Australia) and Goldberg (Washington University, USA) teams utilize conditional and titratable suppression of expression of key PTEX components to provide compelling evidence for the role of the PTEX in export of PEXEL containing and PEXEL-negative exported proteins (PNEPs).

The PTEX translocon contains a heat shock/chaperone protein, HSP101, at the functional heart of the protein complex – here it appears responsible for unfolding the protein to be exported prior to threading through a pore formed from EXP2. Fusing HSP101 to a dihydrofolate reductase destabilization domain (DDD), which is stabilized by trimethoprim (TMP), Beck et al. found that these manipulated parasites require the presence of TMP (equivalent to stable HSP101) to survive, and stage-specific removal of TMP leads to stalled growth at that stage. This applied to both early asexual and early sexual stage parasites, and the effect was reversed by adding back TMP. In ring stages, destabilization of HSP101 blocked export of PEXEL proteins with final destinations in the erythrocyte cytosol and membrane. Importantly, they also show that knockdown of HSP101 function also blocked the export of PNEP – demonstrating the role of the PTEX as a central nexus in the export of apparently all categories of exported proteins from the parasite.
Destabilization of HSP101 at late ring stages afforded a view of proteins exported from this stage onwards. Importantly, export of the PNEP variant antigen/virulence factor P. falciparum erythrocyte membrane protein 1 (PfEMP1) was blocked. It is known that transport of PfEMP1 to the erythrocyte surface requires other exported proteins, so the authors caution that they are unable to unequivocally state that PTEX is directly involved in transport of PfEMP1 itself. The authors also show that the destabilized HSP101 protein has greatly reduced interaction with other members of the PTEX complex (EXP2 and PTEX150), showing that the overall integrity of the PTEX complex is required for efficient function.
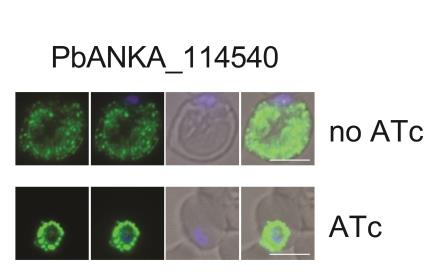
Demonstrating similar results, but in a mouse malaria model, Elsworth et al. conditionally knock down HSP101 expression in P. berghei using an anhydrotetracycline (ATc) regulated transactivator element. P. berghei growth was inhibited, and export of antigens to the infected erythrocyte surface was reduced. Export of two P. berghei PEXEL-containing proteins that normally localize to the erythrocyte cytosol, as well as a specific PNEP that localizes to the erythrocyte membrane, was blocked in HSP101 knockdown parasites. In P. falciparum, they applied the inducible glmS ribozyme system to knock down the expression of PTEX150, a predicted component of the PTEX. The ribozyme degrades tagged mRNA when induced with glucosamine. Titratable PTEX150-knockdown parasites showed varied progression to the early trophozoite stage, but no further. Similarly, PTEX150 disruption blocks export of both PEXEL and PNEP proteins, including PfEMP1.
Together, these studies elegantly demonstrate the application of new molecular tools in exploring the function of PTEX as a translocon mediating the transport of a wide range of proteins central to intraerythrocytic survival and development. The challenge now remains to elucidate the details of the export mechanism and the roles of the additional PTEX accessory proteins. That said, the potential of this nexus for protein export, essential for parasite development and pathology, as a novel drug target would suggest that we will be hearing again from the PTEX community regularly over the coming years!
I read your
blog. Its really good. you have nice contecnt and very good information. Its
very helpful to all.
Glucosamine Exporters, Glucosamine manufacturers
Kindly see my
website also.