
The tapeworm Gyrocotyle discoveryi, an internal parasite of the deep-sea ‘rat fish’ Hydrolagus mirabilis. (Photo credits: R. A. Bray)
Tapeworms are likely to be the oldest recognised type of parasitic worm, with the long, segmented, intestinal-dwelling adult form first described by the ancient Greeks. Like other parasitic flatworms (e.g. flukes) they are highly adapted to an obligatorily parasitic lifestyle and have changed almost beyond recognition from their free-living flatworm cousins.
As adults, they live in the intestines of vertebrates where they absorb food directly through their skin. Thus while flatworms typically have simple, blind guts lacking an anus, tapeworms have dispensed with the gut altogether, letting their host break down nutrients for them.
Meanwhile, each of their segments – numbering 3 to 1000s depending on the species – contains a complete set of male and female reproductive organs (i.e. hermaphrodite). Thus over 95% of the tapeworm adult body is dedicated to the production of eggs! These eggs are eventually released from the intestine into the environment, but to become an adult they need at least one additional host (and sometimes two or three) – the intermediate host – in which the larvae transform into non-segmented juvenile worms, ready to commence sexual development in a vertebrate host and start the cycle again.
Putting the tape in tapeworms
But where did the tapeworm form that we all recognise come from? As it happens, even today not all tapeworms are segmented and it is clear that this adaptation for increasing the number of offspring is a relatively recent feature in their evolution.
Without a fossil record there is effectively no direct evidence of tapeworm ancestry, but we do know from comparative studies of both anatomy and genetic sequences that the ‘true’ tapeworms (the group that includes all segmented species) have two extant (living) evolutionary tapeworm cousins. These two independent lineages each contain only a handful of species which share fundamental underlying characteristics of ‘true’ tapeworms, but otherwise bear little outward resemblance.
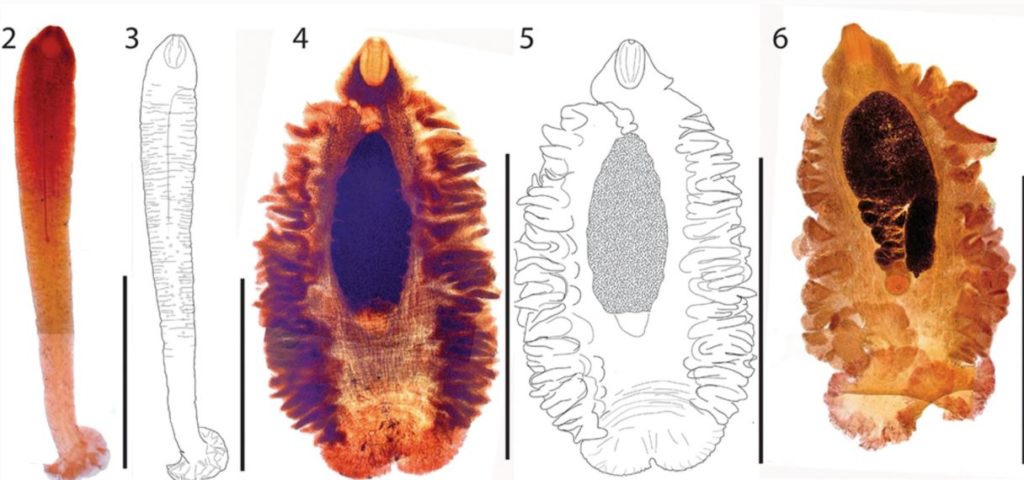
One of these lineages is the Gyrocotylidea, named after their characteristic ‘rosette’ organ (see image at the top). They are exclusively parasites of deep-sea chimaeras, or ratfishes, that together with sharks and rays comprise the cartilaginous fishes. Like true tapeworms, adult gyrocotylideans live in the intestinal tract of their host where they get their nutrition; however, they are not segmented and unlike flatworms generally, are not particularly flat!
Although known since the late 1800s, few new species or other reports on this unusual group have been published in the last 150 years. However, in 2020 we reported on two new species from chimaeras collected in the North Atlantic.
Uncommon parasites that are… uncommonly common
Whilst as a group gyrocotylideans are uncommon compared to the huge diversity of ‘true’ tapeworms (estimated to include more than 20,000 species), as individuals they are uncommonly common in their host group, with the percent of ratfishes infected reaching near 100% in some surveys.
Thus should you have the chance to collect and dissect a ratfish, the chances of finding a gyrocotylidean are high. Unfortunately, sampling from the deep-sea is itself a rare opportunity and of the known ratfish species, most are seldom encountered. The present study was based on collections in the North-East Atlantic made over three years providing the opportunity to investigate the presence of gyrocotylideans in two species of ratfishes never previously examined, revealing two new species of parasite.
Survival of the adequate
Chimaeras are a relictual group of cartilaginous fishes whose diversity is known from the fossil record to have been far greater in the distant past. Although they rarely play host to ‘true’ tapeworms, their evolutionary cousins, the sharks and rays, play host to a significant part of the diversity of true tapeworms. As specialist parasites of chimaeras, gyrocotylideans also appear to be a relictual group, although their past diversity is not likely knowable.
Their contemporary diversity consists of a single known genus (Gyrocotyle) represented by a handful of named species, none of which can be distinguished on the basis of anatomy. Whether the group is really represented by a single, widespread species has long been debated.
The very deep sea is a highly stable environment compared to land – and in the relative absence of strong, dynamic selective forces, ‘survival of the adequate’ may be a better model than ‘survival of the fittest’ in explaining deep sea communities and their potential for relative stasis, compared to communities near the surface and on land.
However, while gyrocotylidean tapeworms are highly uniform in their appearance, the small amount of sequence data that have started to accumulate suggest that there is a significant amount of divergence among parasites sampled from different geographic areas and host species. Accumulating such divergence suggests that the parasites are genetically isolated by geography, host ecology and/or other factors. However, if this were the case, we would expect it to eventually translate into differences in their appearance (i.e. phenotype) as well, as genetic ‘drift’ will occur even in the absence of pressures caused by changes in the environment. Explaining the contemporary diversity of these unusual tapeworms will surely require a better understanding not only of their genetics, but also of the enigmatic ecology of their deep sea hosts.
One of the ten remarkable marine new species from 2020
As for previous years, the World Register of Marine Species (WoRMS) has again released the annual list of the top-ten marine species described by researchers during the year 2020 to coincide with World Taxonomist Appreciation day (March 19th)! Gyrocotyle haffii, or ‘Haffi’s Upside-Down Tapeworm‘, has been chosen among these top ten species. This new species is named in honour of Professor Harford ‘Haffi’ Williams in recognition of his contribution to the understanding of this group of tapeworms.

Comments