
Plants are central to life on Earth: they serve as the foundation for most of its ecosystems, and – at a more fundamental level – generate much of its oxygen. Humans exploit them in agriculture, to source drugs and medicine (not so much any more), and for decoration (still ongoing).
plants have had to evolve regulatory systems to help them survive in an ever-shifting environment
Despite their variety, plants share an obvious drawback: their immobility. As a result, plants have had to evolve regulatory systems to help them survive in an ever-shifting environment that they cannot simply walk away from. Though perennially of interest, with the added pressures of climate change and increasing population, elucidating their mechanisms of survival has never been more important.
The Sainsbury Laboratory Cambridge University opened in 2011, with the goal of creating a collaborative and interdisciplinary research environment through which the underlying regulatory mechanisms of plant growth and development could be clarified. As such, it was a prime venue for this year’s meeting – The Sainsbury Laboratory Symposium 2016 – which brought together an international assortment of plant developmental researchers, studying processes that are inherent, as well as those generated in response to the environment.
Plant sense
A recurrent theme throughout the meeting was plant sensing: how it occurs, and how it drives development and growth. To begin at the beginning (or the end, depending how you look at it), Gwyenth Ingram spoke on how a plant makes seeds, a process that requires coordinated development of several tissues that are essentially isolated from each other, precluding chemical communication. Mechanosensitivity instead drives seed development, the developing seed expanding like, she suggested, a balloon. Last year, her group demonstrated the second cell layer in to be the mechano-responsive region: with changes in microtubule dynamics and thickening of the outer cell wall increasing expression of ELA1 (a regulator of seed size and gibberellin metabolism).
Mechanosensitivity drives seed development, the developing seed expanding like a balloon
For coordination of growth once the plant is out of the seed, one family of plant signaling molecules associated with an ever increasing number of roles is the strigalactone family.
In the opening Keynote talk, Sofie Goormachtig discussed her work on elucidating the underlying strigalactone signalling networks in the root whose crucial role in shaping root architecture is dependent on the hormonal landscape produced from the cross-talk of two other signaling molecules, auxin and cytokinin, so that root architecture is shaped by three tightly integrated signaling networks, enabling the flexibility required for adaptive responses to environmental change.
Predatory Parasites (it’s not all good)
Strigalactone signaling underlies the nodulation response at the early stages of symbiosis with nitrogen-fixing bacteria – but it is also intercepted by less desirable elements of the environment.
Some of the most notorious parasites are those of the genus Striga: “witchweeds”. These parasitic weeds appear from seemingly nowhere, and cause massive crop loss – most often in sub-Saharan Africa, but they have also been seen in other regions. Striga lack the ability to photosynthesize, instead relying on a host that they perceive through strigalactones released into the soil.
Ken Shirasu, interested in establishing the molecular mechanisms of striga-like parasitism has employed Phtheirospermum japonicum as a model organism, which, unlike striga, is a non-obligate parasite – its ability to live outside a host making it easier to study. This has, for instance, enabled him to look more closely at the haustorium – an invasive organ that establishes vascular connections with the host.
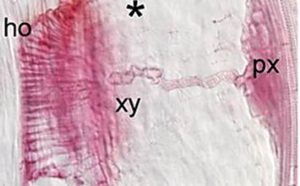
Using forward genetics, his group was able to demonstrate earlier this year that the root hair development program is adapted for haustorial hair formation in P. japonicum, which facilitates transport of nutrients from host to parasite – and they have since shown that development of the haustorium is dependent on the gene YUC3, an auxin biosynthetic gene, with auxin strategically concentrated at the haustorium contact site – another strigalactone-auxin connection.
Many parasites subvert plant growth for their own ends, cyst nematodes providing a particularly intimate example, reported by Melissa Mitchum. In plants, the CLE family of proteins regulate stem cell pools. Cyst nematodes, in an impressive feat of cross-kingdom mimicry, generate their own CLE-like proteins to promote feeding site formation, through manipulation of the plants’ cell proliferation pathways, though they are still dependent on host cellular post-translational machinery to “mature” their CLE-like proteins.
Adapting and – future-proofing?
It is no news that plants adapt their morphology to optimize environmental resource acquisition, though non-invasive imaging approaches, such as X-ray based microCT,used by Malcolm Bennet to study root adaptive responses as they probe the soil for water and nutrients, have shown we didn’t know it all.
Roots, for example, well known to adapt the direction of their growth in response to water level, also adapt the position of lateral roots in response to water level, and to heterogeneous distribution of nutrients.
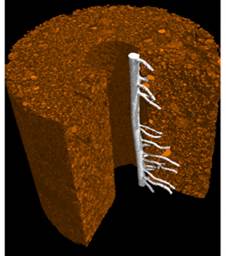
Plants provide many examples of impressive reactive growth adaptations – but how about proactive ones?
These were the topic Jose Gutierrez-Marcos addressed, talking on his work on epigenetics.
He and his colleagues reported earlier this year that high salinity exposure of parent plants confers an improved salinity tolerance on the offspring, and that this increase in robustness is coupled to maternally inherited changes in DNA methylation state, particularly at epigenetically labile regions, capable of spontaneously changing their DNA methylation state.
When stressors are removed, stress-associated methylation states gradually revert. Perfect, it might seem, for rapidly adapting to short-term environmental fluctuations, since stress-associated epigenetic changes are at once stable enough to be heritable over multiple generations, without being irreversible.
Tip of the iceberg
Needless to say, this barely scratches the surface of the astonishing adaptability of plant life, and the growing understanding of environmental effects, internal to the developing plant or external and beyond its control. But advances in understanding of the molecular web underlying plant plasticity will, we must hope, bring plant adaptations increasingly under our control.
Christian Matheou
Latest posts by Christian Matheou (see all)
- Mouse models, tau tangles and neurodegeneration in Alzheimer’s disease - 3rd January 2017
- Rooted in a changing environment - 23rd September 2016
- Learning from plants: new routes to engineering disease resistance - 22nd February 2016
Comments