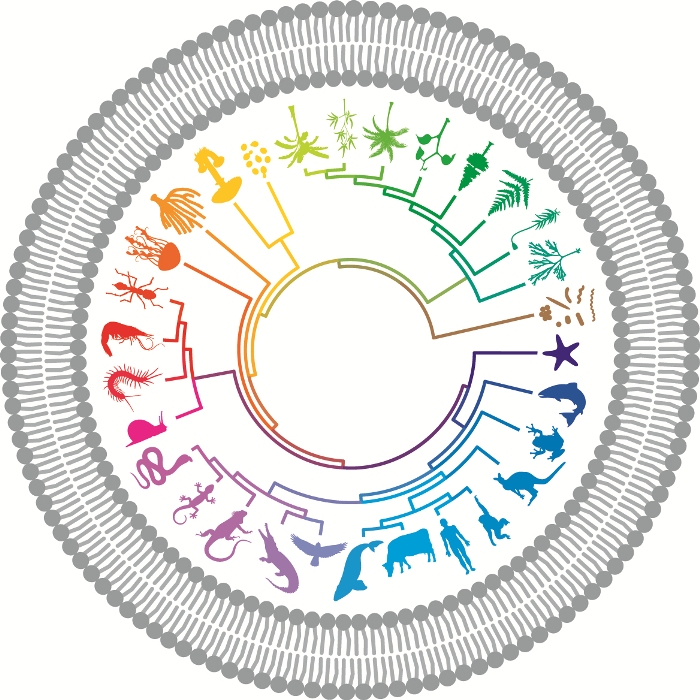
 When BMC Biology launched its iconic image we acknowledged the extreme artistic licence of portraying selected vertebrate phyla pictorially while whole microbial kingdoms were denoted with a single blob. This was not intended to signify a lack of interest in the microbial world on our part, and to update our readers on a major effort to explore its taxonomic diversity and role in the biosphere, we invited the instigators of the Earth Microbiome Project (EMP), launched in 2010 with the aim of sampling the microbial diversity of the planet, to give us a progress report.
When BMC Biology launched its iconic image we acknowledged the extreme artistic licence of portraying selected vertebrate phyla pictorially while whole microbial kingdoms were denoted with a single blob. This was not intended to signify a lack of interest in the microbial world on our part, and to update our readers on a major effort to explore its taxonomic diversity and role in the biosphere, we invited the instigators of the Earth Microbiome Project (EMP), launched in 2010 with the aim of sampling the microbial diversity of the planet, to give us a progress report.
In their short comment article on the achievements and aspirations of the EMP, Jack Gilbert, Janet Jansson and Rob Knight deliver a positive message on crowd-sourcing as an approach for a project of this scale and reach. As of this July they have processed 30,000 samples from a wide variety of ecosystems (including the human microbiome), all collected by collaborators from independent research projects who are willing to make their data available with unrestricted access immediately after analysis.

So far the focus has been on 16S rRNA sequence analysis, and the considerable challenge of developing an open-source analytical platform that not only gives information on the phylogenetic structure of each sample, linked to data on the environment from which it was collected, but which also combines the data from all samples to generate a master table of operational taxonomic units that delineates the abundance of each type of organism in each environment.
This enables a correlative analysis of environmental parameters and microbial community structure and the results are shaping which questions should be addressed and which biomes targeted in selecting the next 20,000 samples to be processed. The original final target of 200,000 samples remains in place, as does the plan to do metagenomic analyses and metabolic modelling as well. Metagenomic analyses have currently been limited to a few sample sets, however, and current efforts with respect to this aspect of the EMP are on developing analysis pipelines to incorporate these data fully into the project.
Readers of a recent comment by Bill Hanage in Nature might ask whether there is a danger of interpreting too much from 16S rRNA data and of overhyping the importance of variations among microbiomes. But while not shy about the ambition of the EMP and the remarkable energy with which it is being pursued, Gilbert, Janssen and Knight do not claim to be saving the world.
They make it clear that the project is amassing correlative data that does not in itself prove causation, and that its value lies largely in co-ordinating their collection, and making them available on an open analytical platform. The hope is, of course that testable hypotheses of cause and effect will emerge (especially as more metagenomic and metabolic data are included), and that it will be possible to cure some ills (both human and environmental) by manipulating the microbiota.
Some hints of this are given in the selected projects discussed briefly in the article, for example the one identifying how loss of microbial diversity and changes in the carbon cycle are linked with agricultural land use in the American prairies. But the issue directly addressed by the project is the primary need to know more about the genetic and functional diversity of microbial communities in different environments. And although 16S rRNA phylogenies are (as Hanage points out) a coarse indicator of diversity, they are a starting point and already the EMP has greatly expanded our knowledge of microbial diversity. Six million genus or species level taxa were identified from the first 10,000 samples, of which only a small fraction could be mapped to previously known 16S rRNA phylogenies.
The distorted underrepresentation of microbes in our BMC Biology icon becomes ever more egregious, and we must once again plead artistic license and hope to continue to avoid an award from Jonathan Eisen for our twisted tree.
Penelope Austin
Latest posts by Penelope Austin (see all)
- Economists listen to Ecologists: On Biology at the World Economic Forum - 9th March 2018
- The secret language of behavior - 31st January 2017
- Restoring a lost microbiome to a model worm - 12th May 2016
No chance. Many many more awards will be coming if you do not fix the tree …